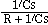 .
.
Such an analysis suggests a theoretical entity, an elementary unit of experience, which corresponds to an activity site (such as a synapse) in the brain. The structure of experience is determined by connections (e.g. dendrites) between these activity sites; the connections correspond to temporal patterns among the elementary units of experience, which can be expressed mathematically. This theoretical framework illuminates several issues, including degrees of consciousness, nonbiological consciousness, sensory inversions, unity of consciousness and the unconscious mind.
First, science is a public enterprise; it seeks knowledge that is independent of the individual investigator by limiting itself to public phenomena. Ultimately it is grounded in shared experiences, for example, when we both look at a thermometer and read the same temperature. Traditionally science has accomplished this by focusing on the more public, objective aspects of phenomena (e.g. temperature as measured by a thermometer), and by ignoring the more private, subjective aspects (how warm it feels to me). In other words, science has restricted itself to facts about which it is easy to reach agreement among a consensus of trained observers. Although this restriction has aided scientific progress, it prevents the scientific study of consciousness, which is essentially private and subjective.2
Second, science's neglect of the subjective is also apparent in its reductive methods. For example, once the experiential phenomenon of temperature has been separated into its subjective and objective parts (felt vs. measured temperature), the objective part can be reduced to other objective phenomena (mean kinetic energy of molecules), but the subjective components of the original phenomenon remain unreduced. Although this approach has been very fruitful for the development of physical theory, it fails when the topic of the investigation is precisely that subjectivity that it ignores.
In summary, the standard reduction pattern in science, which reduces the objective to the objective, cannot solve the hard problem, which deals with the relation between the subjective and the objective. If reduction is to play a role at all, it must take a different form.
Finally, science traditionally seeks facts - observations - that are independent of the observer; this supposes that the observer can be separated from the observed (another aspect of the subject-object distinction). However, in confronting the hard problem we cannot separate the observer and the observed, for consciousness is observation, the subject experiencing the object. That is, experience comprises both observer and observed, the termini of the arrow of consciousness. Separating the two breaks the very connection that we aim to study.
An analogy may make this clear. The aperture of a camera is its `window to the world', since any image in the camera must come through the aperture. (For the sake of the analogy we suppose the camera cannot be opened in any way.) From within the camera the aperture per se is not visible; all we can see is the image it transmits, the scene at which it is aimed. Although the aperture is visible only by virtue of the images it transmits, observation nevertheless shows that certain characteristics of the image (focus, brightness, depth of field) are more a consequence of the aperture than of its content. Thus the aperture may be investigated indirectly. So also we may investigate the structure of consciousness independently of its content.
It may seem that by advocating such private `observation' of consciousness, we have abandoned all hope of publicly validatible science, but it is worth remembering that all observation is ultimately private. Science has developed methods (such as measurement) that, in a context of shared training and experience, lead to general agreement among qualified observers (with varying theoretical commitments), and thus provide a reasonably stable body of public facts, which may be used for the support or critique of theories.3 To bring consciousness into the scope of science will require a body of appropriately trained observers; the public facts necessary for a scientific theory of consciousness will emerge from their consensus.
The camera analogy shows the importance of training, for the relevant phenomena, e.g. depth of field, might not be apparent to untutored observers. The difficulties with `split-brain' and `blind-sight' patients as informants also illustrate the need for trained observers. I believe that the best example of the kind of training required comes from phenomenological philosophy and psychology (see, e.g., Ihde, 1986).
In summary, although consciousness cannot be reduced to physical phenomena by the standard reductive methods of the sciences, it can be investigated to yield publicly validatible facts about the structure of consciousness, which can be related, in turn, to the observations of neuroscience.
The phenomena are the starting point of all science, for they are what is given to us (cf. Latin data = given things).4 However, this is easily misunderstood for, at least since the appearance of logical positivism in the philosophy of science, there has been a tendency to suppose that the phenomena are simple things, such as sense data. `Red-here-now', that is, the current experiencing of a patch of red at a particular location in the visual field, is a well-known example. The phenomenologists, especially Husserl and Heidegger, have demonstrated the incorrectness of this view, for rarely, if ever, do we actually experience red-here-now; they have revealed some of the complexity of real phenomena.
Suppose, for example, you rotate an ordinary die in front of me and ask for a phenomenological account of what I see. I would be incorrect to describe a certain arrangement of black ovals in white parallelograms, both of systematically changing shape. That does not accurately describe the phenomenon as I experience it, for I recognize the object and so it is seen as a die, and I see it rotating in space, not changing shape in some mysterious way. Even if I were unfamiliar with dice, I would see the rotation of a white cube marked with spots. Indeed, it would take very unusual conditions to make me see the die as parallelograms and ovals changing shape. (Such a situation, a consequence of a brain tumor, is described by Oliver Sacks in the title essay of his Man who Mistook his Wife for a Hat.)
An additional complexity of phenomena is that they are not entirely in the here-and-now; for example, they include some foreshadowing of the future. We should be surprised if, as the die rotated, we discovered that an appearing face was unusually marked, or that it was missing and exposed a hollow interior. This shows that the die phenomenon incorporates expectations about its unseen faces.
These expectations go beyond the visual; for example, we also have expectations about the hardness and weight of dice, and if we see two dice in an open palm, we have the expectation of some kind of dice game. All these and more are part of `phenomenal field' surrounding the visual perception of the die. Further, we see that much of the phenomenon is a construct, both of the culture and of individual experience.
In summary, phenomena are not simple; they are highly complex and strongly coupled to the rest of the phenomenal world in which they appear. Therefore some training is necessary to be able to observe phenomena accurately and to analyze the structure of the phenomenal world. Nevertheless I believe that phenomenological training of this kind provides a basis for making the characteristics (though not the experience) of consciousness public.
At the highest level the phenomenal world can be analyzed along modal and functional lines (appearance, sound, smell, memory, intention, etc.), but the rotating-die example shows that these components are far from independent (the rotating die is not visual alone, but includes kinesthetic and other aspects). Fortunately we can expect neuroscientific investigations of functional areas and pathways will correct erroneous preconceived ideas about the structure of the phenomenal world. This analysis of consciousness into components of different kinds can be called a qualitative reduction.
A different kind of reduction, which analyzes some aspect of consciousness into constituents of the same kind, may be called quantitative. This analysis is suggested by topographic maps, which are ubiquitous in the brain. A familiar example is the somatotopic map in the somatosensory cortex: nearby parts of the body are mapped to nearby parts of the cortex, so that the arrangement of neurons mimics the arrangement of the body. Similarly, in the early vision areas we find retinotopic maps, where neurons are arranged in a pattern mimicking the arrangement of their receptive fields in the retina.
The receptive field of a neuron in a topographic map refers to the stimuli to which it responds; for example, a neuron in a somatosensory map might respond to pressure on a particular patch of skin, or a neuron in visual cortex to a particular patch of the retina. In such a case we can identify the subjective experience corresponding to activity in this neuron, namely, pressure in that patch of skin, or light on that patch of retina. I call such a `little bit of experience' is called a phenomeniscon or protophenomenon.5 Further, we can see how, to a first approximation, an objective tactile or visual sensation can be reduced to a large number of receptive fields of this kind. Thus we have a phenomenological subjective-to-subjective reduction (sensation to protophenomena) paralleling the neurological objective-to-objective reduction (topographic map to its neurons, and sensory surface of skin to receptive fields).
This may seem to be a return to the red-here-now model of phenomena, but there are important differences. First, neurons have functional receptive fields that are more abstract than simple spatial patches. For example, in primary visual cortex are neurons whose receptive fields are four-dimensional combinations of retinal location, spatial orientation and spatial frequency (see MacLennan, 1991, for a survey).
Next, as shown by the rotating-die example, there is much more to ordinary phenomena than the sense data, so we will have to take account of nonsensory protophenomena that represent bits of expectations, interpretations, intentions and many more abstract properties. Second, few neurons have simple fixed receptive fields, since even sensory neurons receive inputs from higher brain areas; therefore, virtually all protophenomena depend on other protophenomena. Finally, although I have used a simple sensation as an example, because its protophenomena are easy to visualize, sensation is only one aspect of most phenomena (many of which involve no sensation).
The preceding model can be extended to nonsensory neurons as follows. The activity of a sensory neuron reflects the presence of a stimulus in its receptive field, which is a region of some (possibly abstract) physical space (pressure, light, sound, heat, space, frequency, orientation, etc.); it responds maximally to stimuli in that region. Similarly, the activity of a nonsensory neuron reflects the activities of other neurons, and so it has a functional receptive field, comprising certain patterns of activities of its input neurons, to which it responds. That is, as sensory neurons respond to combinations of physical energy, so nonsensory neurons respond to combinations of neural activity. Turning to the subjective side, we see that, as a sensory protophenomenon corresponds to the physical phenomena in a sensory neuron's receptive field, so a nonsensory protophenomenon corresponds to the protophenomenal intensities in a nonsensory neuron's functional receptive field.6
We have seen that activity in some component of the nervous systems reflects the extent to which its functional receptive field is occupied at that point in time. Subjectively, this activity corresponds to the intensity in consciousness of a protophenomenon corresponding to the site of that activity. Therefore I hypothesize a one-to-one relationship between protophenomena and certain activity sites in the brain, and further hypothesize that the intensity of a protophenomenon varies directly with the neurological activity at that site. What are these activity sites?
Following Sherrington, who said, `Reflex action and mind seem almost mutually exclusive - the more reflex the reflex, the less does mind accompany it', Pribram has argued that consciousness is associated with graded dendritic processes rather than all-or-nothing axonal spiking (Miller, Galanter & Pribram, 1979, pp. 23-4; Pribram, 1979, pp. 104-5; Pribram, 1991, 7-8). For concreteness I will accept this hypothesis and take synapses to be the activity sites (though the identification is not crucial to most of the following).7 Candidates for the activity of the synapse include presynaptic membrane potential, postsynaptic membrane potential and neurotransmitter flux across the synaptic cleft. Since for the most part each is proportional to the others, it doesn't matter much which we pick; for concreteness, I'll hypothesize postsynaptic membrane potential.
The easiest way to understand protophenomena is to think of them as the atoms (indivisible constituents) of consciousness. As atoms make up macroscopic objects (trees, tables, etc.), so protophenomena make up phenomena. In both cases, the effect of each individual element on the whole is usually minute. Indeed, individual protophenomena, like atoms, are invisible but, just as atoms combine into visible objects, so protophenomena combine into phenomena.
On this view, the state of the phenomenal world, that is, the content of consciousness, is identical with the intensities of all the protophenomena. The appearance of coherent or stable phenomena can be identified with cohesive or coherent patterns of intensity among the protophenomena (just as macroscopic objects and events can be identified with cohesive and coherent patterns of activity among atoms).
Here again the atomic analogy is helpful. When atoms were first postulated, they were theoretical entities; indeed it is only in recent years that they have become directly observable (still, of course, through instruments). At first many respected scientists denied their existence, while admitting their convenience for theory. In time, their explanatory value became so great that they were accepted as real.
Now we accept atoms (or more elementary particles) as the ultimate constituents of matter, which cause the properties of macroscopic objects, and in this sense, in physical theory, atoms are prior to trees. Nevertheless, in experience, trees are prior to atoms. Similarly, in phenomenology, protophenomena are prior to phenomena, for they are the causes of phenomena, but in experience phenomena are prior to protophenomena; we experience trees, and by analysis break them down into protophenomena.
Another ontological issue is whether an isolated synapse (in a petri dish or simple organism, for example) has an associated protophenomenon. One is that protophenomena, as components of the phenomenal world, make sense (`exist') only in the context of a sufficiently complex nervous system. (`Sufficiently complex' is of course a matter of degree.) Such `emergent existence' is not uncommon in scientific theories. For example, sound is a compression wave in air or another medium. The theory assigns a pressure to every point in the medium, yet it makes little sense to talk about the pressure (or sound) of a single air molecule. The compression wave, which comprises elementary units of pressure assigned to individual molecules, makes sense only in the context of a large number of molecules. Similarly, I think it may make sense to assign protophenomena to activity sites only in the context of a large number of activity sites. This emergence does not make the protophenomena any less real; they are as real as the elementary units of pressure which constitute the sound wave.
I have said that conscious states are the totality of protophenomenal intensities, so phenomena, as aspects of the phenomenal world, are cohesive and coherent patterns of protophenomenal intensity. I see no reason to hypostasize these patterns by postulating (subjective) entities corresponding to phenomena (images, ideas, perceptions, etc.). The coherence of the intensity patterns constitutes their appearance in experience (as will be explained in more detail later). By analogy, to explain the coherent physical effect of a baseball on a window it is not necessary to postulate the existence of anything beyond the baseball's constituent atoms, such as a `ball entity', to represent the ball's coherence. So also, the collective action of protophenomena are sufficient to explain the experience of a coherent phenomenon.
Second, I hypothesize that neural differences imply phenomenal differences. Here the reason is Occam's Law, for we would otherwise have to suppose that some activity sites (e.g. synapses) have associated phenomenisca while others don't. Although this may be the case, I see no evidence supporting it.8
Our discussion of topographic maps suggests that they have an important role to play, but how, precisely, do the spatial relations among neurons lead to phenomenal (spatial and more abstract) relations among protophenomena? Although there may be some diffuse electrical and chemical effects on the activity of neurons, it seems that in general the spatial arrangement of neurons is significant only because it correlates with connectivity: nearer neurons are more likely to be connected than are more distant ones, and connections create dependencies between neurons.
If the protophenomena were completely independent of each other we would not have a phenomenal world; we would have chaos rather than a cosmos. Conversely, the structure of the phenomenal world implies necessary correlations among the intensities of the protophenomena. By looking at their neurological correspondents, we can see that these necessary correlations come about through physiological processes and neural connections. The connections define the possible states of consciousness and thus the phenomenal world. What may be called contingent correlations arise from outside the nervous system, but they are not part of the structure of the phenomenal world unless they affect neural connectivity. For example, one neuron forms a synapse on another, so that the pre- and postsynaptic activities are correlated (sequential correlation). A special case is where there are divergent connections (one neuron synapsing on two others), which create parallel correlation among postsynaptic activities.
These necessary correlations reflect the structure of the phenomenal world in that occurrence of a protophenomenon makes more (or less) likely the occurrence of other protophenomena. It is these relations between protophenomena that gives them their meaning. By analogy, a set of pixels constitutes a picture only when combined in a certain arrangement (relations of nearness or adjacency); with a different arrangement they would be a different picture; so also, with different relations, a set of protophenomenal intensities would constitute a different conscious state.
Let's consider protophenomenal relations in more detail. Neurologically, the activity at a synapse is a complex spatiotemporal integration of the activities of the synapses to which it is connected. To a first approximation this process is linear, and can be described by the methods of linear system analysis (MacLennan, 1993b), which shows there is a certain spatiotemporal pattern to which the synapse shows the maximum response. Indeed, this pattern can be used to describe the temporal response of the synapse to any spatiotemporal signal, in so far as the synapse behaves linearly. For this reason I will call this maximum-response spatiotemporal pattern the characteristic pattern of the synapse.9 (Technically, the response of a synapse is a convolution of its characteristic pattern with the input signal.)
This account may be transferred directly to the phenomenal realm. Each protophenomenon has a characteristic pattern, which is the spatiotemporal pattern of intensities of its input protophenomena that will maximize its intensity. Further, its characteristic pattern determines (by convolution) the protophenomenon's time-varying intensity in response to any spatiotemporal pattern in its input protophenomena. As a consequence we can give a mathematical theory of the causal relations among protophenomena (see the Appendix to this paper). The characteristic patterns may be simple, as when a protophenomenon corresponds to a conjunction or disjunction of protophenomena, or more complex, as when they respond to appearance or disappearance of protophenomena, rhythmic or other temporal patterns in protophenomena, priming or inhibition of future occurrences of protophenomena, etc.
Overall, the causal relations among protophenomena are nondeterministic. First consider a synapse formed by a sensory neuron, the activity of which depends on physical stimuli as well as the activities of other synapses. Corresponding to this activity site in the phenomenal realm we have a protophenomenon whose intensity depends on physical processes as well as other protophenomena. But physical processes are not part of the phenomenal world, so such a protophenomenon is phenomenologically undetermined (i.e. not fully determined by other protophenomena); the physical inputs act as independent variables in the phenomenal world. In terms of the ontology of the phenomenal world, they are causal primaries, which does not imply, however, that there are not corresponding phenomenal expectations (as the rotating-die example shows). Thus sensory protophenomena are inherently nondeterministic (i.e. undetermined by the phenomenal world).
Since nonsensory protophenomena depend only on other protophenomena, to a first approximation they can be considered deterministic; indeed their response is defined by their characteristic pattern. This is only an approximation because even nonsensory neurons depend on non-neural processes, such as the physiology of the brain, and the physical environment of the body. Although these effects can sometimes be treated as extra, hidden inputs to the synapses, they are often nonlinear and comparatively nonspecific in their effects, so it is usually better to treat them as phenomenologically undetermined alteration of the characteristic patterns of the affected protophenomena.10
I have described the protophenomenal dependencies from a mechanistic perspective; now it is worthwhile to say a few words from a functional perspective. Topographic maps show us how receptive fields are ordered in space, frequency, speed, color, and many other dimensions, and hence how their protophenomena are ordered in corresponding subjective domains. Thus causal dependencies among protophenomena correspond to order in a variety of dimensions. This order means that objects extended in space or other dimensions will lead to high intensities among closely dependent protophenomena (which will therefore cohere as full-fledged phenomena).
Furthermore, since change is generally continuous, or if discontinuous in some dimensions, then continuous in others, it follows that changing objects tend to move from the receptive fields they occupy to others that are adjacent along one or more dimensions. Think of a visual image of a moving object: it moves between adjacent spatial receptive field; further, its edges change orientation continuously, and the light it reflects changes continuously, and so it also moves from receptive field to receptive field in these dimensions. Phenomenologically, we can say that change tends to be between protophenomena that are strongly connected. Conversely, the presence (high intensity) of a protophenomenon is correlated with the future presence of the other protophenomena that depend on it. In approximate terms, the causal dependencies among protophenomena correspond to the likelihood (or unlikelihood) of change between protophenomena. More accurately, the characteristic pattern of a protophenomenon represents likely antecedent spatiotemporal patterns of protophenomena.
Second, although the adult brain does not generate new neurons, it does generate new synapses for a number of reasons, including injury and learning (Shepherd, 1994, pp. 222-3). Since I have hypothesized that protophenomena correspond one-to-one with synapses, the generation of new synapses implies the generation of new protophenomena, that is, new degrees of freedom in the phenomenal world - literally, `expanded consciousness'. Thus we see that the phenomenal world has a flexible ontology at both the macroscopic (phenomenal) and microscopic (protophenomenal) levels.
It is possible, at least in principle, to attack this problem empirically. We would have to identify some observable protophenomenon, the presence or absence (high or low intensity) of which can be reported reliably by a trained observer, and for which the corresponding synapse (or other activity site) can be identified and made accessible (e.g. through brain surgery). With care we may control some of the variables (e.g. postsynaptic potential) independently of the others (e.g. presynaptic potential), and thus determine which affect protophenomenal intensity. Indeed, one could replace the synapse by devices that are functionally equivalent in one way or another (e.g. electrically or chemically), to determine which are necessary or sufficient for the existence of the protophenomenon.
It will be objected that the investigation depends on subject report, which is a form of behavior, and therefore need not reflect subjective experience. That is correct. Since subjectivity is private, the only way such doubts can be eliminated is for the doubter to be the subject of the experiment.13 Practically, though, the observations would become public through a consensus of trained observers of differing commitments.
From such a demonstration of protophenomena associated with nonsynaptic or even nonbiological objects we could reasonably conclude that a phenomenal world, and therefore consciousness, would emerge from sufficiently complex interconnections of those objects. Nevertheless, my guess is that we will find that the representation and information processing properties of synapses are all that matters, and that other physical systems (such as massively parallel analog computers) with the same capabilities will have protophenomena and, if appropriately structured, be conscious.14
Consider first a hypothetical inversion of loud and soft sounds: Is is possible that I hear as loud what you hear as soft, and vice versa? We can see that this is impossible, since the two ends of the loudness scale have different properties; in particular, as loudness decreases into silence, all pitches become the same, but pitches retain their identity as loudness increases. Not only can we not hear different pitches of silence, we cannot even imagine them. Apparently our auditory systems do not have a separate representation for `silent middle C'; we can put the words together, but we cannot imagine the sound. In other words, there is a `degeneracy' or `singularity' at zero-amplitude, where all pitches collapse together. As Francis Bacon said (Essays, 3), `All colours will agree in the dark'; likewise, in silence all pitches are the same.
Next consider a pitch inversion, in which the sensation of high and low are reversed. This is also impossible, because of the unique characteristics of low pitches: for if we listen to a sine wave of decreasing pitch, our perception of it will change gradually from a tone, to a buzz, to a rhythm. Neurologically, a pitch, which is mapped spatially in the auditory cortex, changes to an amplitude variation, which is mapped temporally. As the pitch decreases below about 1000 Hz., the nerve impulses begin to synchronize with the sound vibrations; below about 20 Hz., they are not perceived as pitch, but as periodic loudness variations (rhythm). In other words, at low frequencies the pitch and loudness axes are not independent; this does not happen at high frequencies, so the low end is topologically distinct from the high. The proposed inversion is impossible.
Finally, we may ask whether the pitch and loudness axes could be exchanged, so that I experience as pitch what you experience as loudness, and vice versa, but this is also impossible, because the two interact in an asymmetric way: low pitches blend into loudness variations, but soft sounds do not blend into pitch variations.
What may we conclude from the impossibility of these inversions? First, that subjective experience of sound must be just the way it is. For example the hearing of a low pitch is identical to intensification of certain pitch protophenomena that are connected in a certain way with loudness protophenomena. This view may seem tautologous, and therefore useless, but it is not. For example, if we discovered an organism with sense organs sensitive to vibrations of another kind (electrical, say), but of similar frequency, so that the similar interrelations hold among the frequency and amplitude protophenomena, we could reasonably conclude that its experience of those sensations would be like sound.
An analogy may clarify this. A picture is an emergent effect of its individual pixels and their relative positions. If we cut a picture in half, it becomes two pictures, because there is no longer a fixed relations between pixels in one half and those in the other. However, instead of cutting the picture, we may gradually separate it into two parts, pixel by pixel, by stretching and eventually breaking the connections between them. The gradual uncoupling of the pixels in the two halves causes the picture to change gradually from one to two. So also, consciousness is emergent from the individual protophenomena and the dependencies between them. As the neural connections are weakened or broken, the protophenomena in the two subworlds decouple from each other, and the one mind becomes two.
This thought experiment demonstrates that the unity of consciousness is a matter of degree. Indeed, in principle we can measure the unity of consciousness by the tightness of the coupling between its protophenomena, for it is this coupling that gives the phenomenal world its coherence. (The tightness of coupling can, in principle, be calculated from the characteristic patterns.)
One may wonder what sort of coupling is sufficient to unify consciousness. For example, in split-brain patients it has been observed that one hemisphere may communicate with the other through transactions with the external world, for example, twitching the skin on one side of the face so that it can be felt on the other. (The patient is unconscious of doing this.) Can `external transactions' such as these effect the coupling of phenomenisca? If so, then our individual phenomenal worlds may not be so independent as we commonly suppose, for any sort of communication couples protophenomena in one mind to those in another. I think the answer is, again, a matter of degree. There is an enormous difference between the bandwidth of the corpus collosum (approximately 800 million nerve fibers) and the narrow bandwidth of most external media. Nevertheless, the interconnection of phenomenal worlds by nonneural physical processes is an thought-provoking possibility.
(1) Unconscious processes may correspond to low-intensity, loosely-coupled protophenomena. By becoming coherent they come into consciousness. That is, unconscious processes are incoherent patterns in protophenomenal intensity. An analogy may clarify this. Project a slide on a screen, and defocus the lens. All of the same information is being projected on the screen as before, but now it is incoherent and the pattern is not salient; this is analogous to unconscious patterns in the protophenomena: they are there but not manifest. Focusing the lens makes the image manifest, which is analogous to the emergence of the unconscious content into conscious experience.
(2) The split-brain operations suggest another solution: in many cases the right hemisphere is unable to respond verbally to problems, and so it cannot express its consciousness. Further, since the consciousness of the right hemisphere is largely disjoint from that of the right, the right forms a kind of unconscious mind for the left. Of course, the right hemisphere is as conscious as the left, and can manifest its consciousness in other ways, but its experience is not part of the left hemisphere's experience (or vice versa). The analogy becomes more striking when we recall that in these patients the hemispheres are not completely disconnected, so the right hemisphere can inject ideas into the left via the brainstem on external transactions. Indeed, split-brain patients experience these communications as inexplicable `hunches' - just like those from the unconscious (Gregory, 1987, p. 743).17 In summary what I, the perceiving-acting-speaking self, experience as the `unconscious mind' may be a equally conscious but loosely coupled part of my phenomenal world, which manifests itself only through hunches, dreams, urges, etc.
(3) Finally, according to the hypothesis of Sherrington and Pribram, discussed earlier, consciousness is associated with graded dendritic microprocesses but not with all-or-none impulses in the axon. Therefore the unconscious mind may reside in the axons, which would make it comprise the more reflexive, instinctive aspects of the psyche. In fact, it is reasonable to suppose that all three of these processes contribute to the unconscious mind (which is primarily, it must be noted, a negative concept).
There are some superficial similarities between protophenomena and the psychons proposed by Sir John Eccles (1990, 1993); they are both elementary units of consciousness associated with synaptic activity in dendrites. The first difference is one of scale: Eccles associates psychons with dendrons, bundles of the apical dendrites of approximately one hundred pyramidal cells. Therefore, a dendron contains approximately 100,000 synapses, and so we could say that a psychon corresponds to approximately 100,000 protophenomena. The second difference is ontological, for Eccles' theory is explicitly dualistic. He takes a psychon to be a causal primary, which can influence synaptic processes by momentarily altering the quantum mechanical probability of an exocytosis of neurotransmitter into the synaptic cleft. In contrast, my theory is monistic, for it views the phenomenal and the neural as two aspects of the same reality; in this sense my view is more akin to Sperry's non-dualist mentalism (Sperry, 1983, ch. 6).
On the other hand, Sperry's theory of emergent causation does not seem to adequately distinguish awareness and consciousness. Sperry (1983, p. 92) says, `Once generated from neural events, the higher order mental patterns and programs have their own subjective qualities and progress, operate and interact by their own causal laws and principles which are different from, and cannot be reduced to those of neurophysiology0.' The focus of emergent causation is on the functional role of conscious phenomena, and so he is dealing with awareness rather than consciousness; he holds `subjective mental phenomena to be primary, causally potent realities as they are experienced subjectively' (p. 79), and speaks of the distinction between `the causal potency of mental experience per se and that of its neural correlates' (p. 91), but does not address the emergence of subjective experience from smaller elements of subjectivity.
The present theory is dualistic in the sense that certain objects in certain situations (namely, activity sites in a functioning brain) have fundamental properties (protophenomena and their intensities), which are not reducible to physical properties. It is also dualistic in that the inherently private fact of experience is not reducible to the phenomena experienced, which are all potentially public (up to a consensus of trained observers). Nevertheless, it is a kind of monism in postulating one `stuff', which happens to have two fundamental, mutually irreducible aspects (phenomenal and physical).
Irreducibility enters in another way, for emergent causation operates in both the phenomenological (mental) and neurological (physical) realms: macroscopic consciousness governs microscopic protophenomenal dynamics (without violating the microscopic protophenomenal laws), as macroscopic awareness governs microscopic neurodynamics (without violating microscopic neurophysiology). (See also Sperry, 1983, pp. 93-6.) Once the philosophical arguments for irreducibility are granted, scientific investigation can proceed by parallel analyses in the phenomenological and neurological realms, each supplying the other with hypotheses, theories and empirical data. However, phenomenologically trained observers will be needed to obtain repeatable observations of the characteristics of consciousness.
As a first step I have proposed a theoretical entity, the protophenomenon, as an elementary unit of consciousness associated with microscopic activity sites in the brain, tentatively identified with the synapses. Like other theoretical entities in science, protophenomena are validated by their explanatory value and their fruitfulness for further progress. According to this theory the phenomenal world results from causal dependencies among the protophenomena, which parallel to the neurodynamical dependencies among the corresponding activity sites; indeed they are described by the same mathematical laws.
Chalmers, David J. (in press), `Facing Up to the Problem of Consciousness', Journal of Consciousness Studies.
Dennett, Daniel C. (1991), Consciousness Explained (Boston, Toronto and London: Little, Brown and Company).
Eccles, J. C. (1990), `A Unitary Hypothesis of Mind-brain-interaction in the Cerebral Cortex', Proceedings Royal Society London B, 240, pp. 433-451.
Eccles, J. C. (1993), `Evolution of Complexity of the Brain with the Emergence of Consciousness', in Rethinking Neural Networks: Quantum Fields and Biological Data, ed. K. H. Pribram (Hillsdale: Lawrence Erlbaum).
Fleck, Ludwik (1979), Genesis and Development of a Scientific Fact, transl. by Fred Bradley & Thaddeus J. Trenn (Chicago and London: University of Chicago Press).
Gregory, Richard L. (ed.) (1987), The Oxford Companion to the Mind (Oxford and New York: Oxford University Press).
Hameroff, S. R. (1994), `Quantum Coherence in Microtubules: A Neural Basis for Emergent Consciousness?', Journal of Consciousness Studies, 1(1), pp. 91-118.
Hempel, Carl G. (1965), Aspects of Scientific Explanation and Other Essays in the Philosophy of Science (New York: The Free Press).
Ihde, Don (1986), Experimental Phenomenology: An Introduction (Albany: State University of New York Press).
Jaynes, Julian (1976), The Origin of Consciousness in the Breakdown of the Bicameral Mind (Boston: Houghton Mifflin).
MacLennan, Bruce J. (1991), `Gabor Representations of Spatiotemporal Visual Images', Technical Report CS-91-144 (Knoxville: University of Tennessee Computer Science Department); accessible via URL http://www.cs.utk.edu/~mclennan.
MacLennan, Bruce J. (1993a), `Grounding Analog Computers', Think, 2, pp. 48-51.
MacLennan, Bruce J. (1993b), `Information Processing in the Dendritic Net', in Rethinking Neural Networks: Quantum Fields and Biological Data, ed. K. H. Pribram (Hillsdale: Lawrence Erlbaum).
MacLennan, Bruce J. (1994), `"Words Lie in Our Way"', Minds and Machines, 4(4), pp. 421-37.
MacLennan, Bruce J. (in press), `The Investigation of Consciousness through Phenomenology and Neuroscience', in Scale and Conscious Experience, ed. K. H. Pribram (Hillsdale: Lawrence Erlbaum).
Maxwell, Grover (1980), `The Ontological Status of Theoretical Entities', in Introductory Readings in the Philosophy of Science, ed. by E. D. Klemke, Robert Hollinger & A. David Kline (Buffalo: Prometheus Books).
Miller, George A., Galanter, Eugene, & Pribram, Karl H. (1979), Plans and Structure of Behavior (New York: Adams-Bannister-Cox).
Pribram, Karl H. (1971), Languages of the Brain: Experimental Paradoxes and Principles in Neuropsychology (Englewood Cliffs: Prentice-Hall).
Pribram, Karl H. (1991), Brain and Perception: Holonomy and Structure in Figural Processing (Hillsdale: Lawrence Erlbaum).
Sacks, Oliver (1985), The Man who Mistook his Wife for a Hat (London: Duckworth).
Searle, John (1992), Rediscovery of the Mind (Cambridge: MIT Press).
Shepherd, Gordon M. (1994), Neurobiology, 3rd ed. (New York and Oxford: Oxford University Press).
Sperry, Roger (1983), Science and Moral Priority: Merging Mind, Brain, and Human Values (New York: Columbia University Press).
Storr, Anthony (1983), The Essential Jung (Princeton: Princeton University Press).
In terms of its electrical activity, a synapse is a voltage-controlled voltage source. On the presynaptic side it exhibits both resistance and capacitance, so we can treat a synapse as the equivalent circuit (Fig. 1). The dendrites, which connect many synapses, have resistance and capacitance, which depend on the diameter of the dendrite, among other things. As is commonly done, I will treat the dendrite as a tree of cylindrical segments of constant diameter.18
The dendritic membrane can be treated as a simple passive circuit, as shown in Fig. 2. Its impedance is Z = R + 1/Cs, since 1/Cs is the impedance of the membrane capacitance. Thus the transfer function of the potential is given by
H(s) = .
Multiplying above and below by C and recalling that t = RC is the time constant of the R-C circuit yields
H(s) =  .
.
The corresponding characteristic function (impulse response) is
h(t) = e-t/t / t.
This decaying exponential smooths any impulse traversing the dendrite.
Consider now the effect of postsynaptic membrane potential on dendritic membrane potential. Figure 3 shows the dendritic connections and the equivalent circuit. The potential eO at the root of the dendritic spine is an effect of the postsynaptic potential eS, acting through the spine neck, interacting with the dendritic membrane potentials on both sides (eL and eR). By applying Kirkhoff's laws we can determine eO in terms of the dendritic conductances (GL, GS and GR) and the membrane capacitance C near the spine. The dependence is easiest to express in terms of the Laplace transforms of the quantities:
EO = 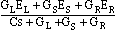 .
.
Dividing above and below by C, and recognizing that Gx/C = 1/CRx = 1/tx (x = L, S, R), the inverse time constant of the dendrite, we have:
EO =  .
.
Let u = 1/tL + 1/tS + 1/tR be the sum of the inverse time constants, and define the transfer function H(s) = 1/s+u. Then the Laplace transform of the potential is
EO = H(s) (EL/tL + ES/tS + ER/tR).
The impulse function corresponding to H is h(t) = e-t/u. The output potential is then given by a convolution:
eO(t) = e-t/u * [eL(t)/tL + eS(t)/tS + eR(t)/tR].
The weighted sum of the potentials is smoothed by the exponentially decreasing impulse function, a consequence of the membrane capacitance. This dependence between the potentials will be depicted by a symbol such as that in Fig. 4.
The presynaptic membrane potential is similar and in fact simpler:
eI(t) = e-t/u * [eL(t)/tL + eR(t)/tR],
where u = 1/tL + 1/tR. To a first approximation, the relation between the pre- and postsynaptic potentials is a simple proportion, eO = ceI. (To be more accurate we would have to consider the diffusion of the neurotransmitter, which would also have a smoothing effect.) I will depict this relationship by the symbol in Fig. 5. The characteristic pattern then is a vector function displaying the signals (eL, eL) to which the synapses is tuned:
hI(t) = h(t) (1/tL, 1/tR) = (e-t/u/tL, e-t/u/tR).
We now have formulas relating the activities at activity sites to each other, and we can reinterpret them as formulas relating the intensities of protophenomena. A diagram such as Fig. 6 shows how the protophenomenal intensities depend on each other, and allow their calculation, at least in principle. In general we can see that the (time-varying) intensity of a protophenomenon will be a complex function of the (time-varying) intensities of those on which it depends, as their intensities may be of its. The products of the transfer functions along each path determine the transfer function of the synapse, and hence its characteristic pattern.
2It should be apparent that I am using subjective and objective to distinguish private, `first person' phenomena from public, `third person' phenomena. As Searle (1992) observes, progress on the mind-body problem has been impeded by the connotations acquired by these terms, viz., the objective is unbiased and factual, whereas the sub jective is biased or distorted. Indeed, I will argue the possibility of unbiased, factual statements about subjective (private, first person) phenomena.
3One cannot ignore the importance of training, shared experience and institutions in the creation of `facts'. Even something so simple as accurately reading a thermometer requires training and skill (e.g. reading the top or bottom of the meniscus). Training is all the more necessary for reading bubble-chamber images and gas chromatographs. The histories of N-rays and polywater show how competent observers can disagree over even the existence of a phenomenon (let alone its measurement). See Fleck (1979) for an informative case study.
4This is true of the empirical sciences, but also of the so-called apriori sciences, such as mathematics, which start from apparently invariable structure of the phenomenal world.
5`Phenomeniscon' (accent on penult), a diminutive of `phenomenon', is used in MacLennan (in press). I am grate ful to David Chalmers for suggesting the alternative `protophenomenon'.
6This description is simplified for the sake of exposition, since sensory neurons also respond to other neurons, and so their complete receptive field includes the activities of other neurons. Correspondingly, the subjective intensity of a sensory protophenomenon depends on the subjective intensities of other protophenomena as well as on objec tive physical processes.
7In particular, my proposal does not exclude the possibility that microtubules, as proposed by Hameroff (1994), are among the activity sites.
8Note, that the claim is only that differences of activity lead to differences of protophenomenal intensity and hence conscious state, not necessarily that the difference will be attended or have a significant effect on future conscious states. Analogously, changing a pixel changes the picture, but we wouldn't ordinarily notice the change. The issue of the unconscious is treated later; suffice it to say that it does not contradict the hypothesized one-to-one relation.
9In physics and engineering it is commonly called the impulse response of the system; it corresponds (via the La place transform) to the transfer function, which describes the system dynamics in the frequency domain.
10Thus there may be phenomenologically causeless change to the phenomenal world; an extreme example is a stroke.
11My concern here is not so much plasticity in the developing animal as plasticity in the adult.
12Chalmers (in press) tentatively reaches the same conclusion on the basis of his `double-aspect principle'.
13If this seems far-fetched, it is worth noting that William McDougall requested, if he should become incurably ill, that Sherrington would perform a cerebral commissurotomy (split-brain operation) on him, so that he might direct ly experience its effect on his consciousness (Gregory, 1987, p. 741).
14It will be apparent from this that I do not accept Searle's reply to the Virtual Minds version of the System Reply to the Chinese Room Argument (MacLennan, 1993a, 1994). See also the thought experiment in Chalmers (in press).
15Thus I disagree which Chalmers (in press) when he asserts, `There are properties of experience, such as the in trinsic nature of the sensation of red, that cannot be fully captured in a structural description'. I will argue that the experience is exhausted by its structure.
16There are a number of definitions of the unconscious; for my purposes Jung's is as good as any: `Everything of which I know, but of which I am not at the moment thinking; everything of which I was once conscious but have now forgotten; everything perceived by my senses, but not noted by my conscious mind; everything which, invol untarily and without paying attention to it, I feel, think, remember, want, and do; all future things that are taking shape in me and will sometime come into consciousness: all this is the content of the unconscious.' (CW 8, ¶ 382; Storr, p. 425)
17This experiencing of interhemispheric communication as communications from an external source (`the gods') is of course a major premise of Jaynes' (1976) theory of the development of consciousness.
18Since we are primarily concerned with dendritic interactions (as the substrate of consciousness), I will not ad dress the soma or axons here.